
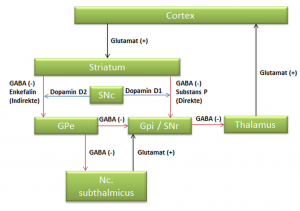
I det indre af hemisfærerne findes en større masse af grå substans. De største af disse kaldes basalganglier (Corpora striata), som er vigtig for bl.a. bevægelse.
Basalganglierne forbinder flere af de store kerner i hemisfærene. Striatum består af putamen og nucleus caudatus og modtager vigtige forbindelser fra hjernebarken. Striatum sender signaler videre til globus pallidus og substantia nigra (den sorte substans). Fra disse går signaler videre til hjernestammen og tilbage til hjernebarken via thalamus. Nc. subthalmicus hører også til basalganglierne.
Ved at påvirke motoriske neurongrupper i frontallapperne og hjernestammen bidrager basalganglierne til kontrol af bevægelser uden selv at sætte dem igang. De har også forbinder med andre dele af frontallappen der har med kognitivefunktioner at gøre.
Basalganglierne ses i et horisontalsnit af hjernen til at bestå af to delvist adskilte dele – en stor del linsekernen (Nc. lentiformis) der ligger lateralt for capsula interna og en noget mindre del halekernen(Nc. caudatus) der ligger medialt for capsula internaog foran thalamus.
Anatomiske opbygning
Basalganglierne består af mange anatomiske strukturer og man kan gruppere dem på flere måder. Generelt inkluderes følgende strukturer til basalganglierne:
- Nucleus Caudatus
- Putamen
- Globus Pallidus
- Substantia Nigra
- Nucleus subthalmicus
Nucleus Caudatus og Putamen kaldes samlet for Striatum eller Neostriatum. Striatum sammen med Globus Pallidus kaldes for Corpus striatum. Putamen og Globus Pallidus kaldes tilsammen for Nucleus Lentiformis (Lentiformis = linseformet).
- Striatum = Nc. caudatus + Putamen
- Corpus striatum = Nc. caudatus + Putamen + Globus Pallidus
- Nc. Lentiformis = Putamen + Globus Pallidus
Neurotransmittere
I basalgangliernes kredsløb er det især to neurotransmittere der er vigtige. Det drejer sig om Glutamat der er excitatorisk og GABA der er inhibitorisk. Kredsløbet består ofte af en serie af excitatoriske og inhibitoriske neuroner som tilsammen bliver enten excitatorisk eller inhibitorisk.
Nogle gange kan det være svært at overskue om summen af disse excitatoriske (+) og inhibitoriske (-) neuroner bliver enten excitatorisk (+) eller inhibitorisk (-). Men her kan man faktisk anvende en af matematikens regneregler der siger:
- + gange + = +
- – gange – = +
- + gange – = –
- – gange + = –
Fordi hvis man har to excitatoriske neuroner efter hinanden er resultatet excitatorisk. Har man to inhibitoriske neuroner efter hinden er resultatet også excitatorisk. Har man en excitatorisk neuron efterfulgt af en inhibitorisk neuron eller omvendt er resultatet inhibitorisk. På samme måde kan man altså gange flere neuroner sammen, f.eks. giver “+” gange “-” gange “-” resultatet “+”.
Udover GABA og Glutamat anvendes også Enkefalin og Substans P som neurotransmitter sammen med GABA.
Direkte og indirekte kredsløb
Der er både et direkte og et indirekte kredsløb som forbinder cortex med basalganglierne og herefter med thalamus og cortex igen. Justeringen mellem disse gør at man kan udføre en bestemt motoropgave uden at blive forstyrret af andre motoropgaver samtidig.
Nedenstående figur præsenterer både det direkte og det indirekte kredsløb.
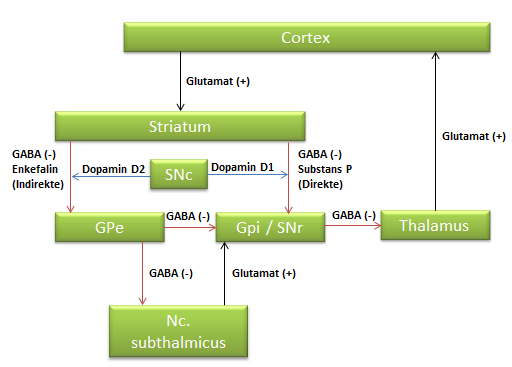
Det direkte kredsløb går fra Cortex til Striatum (via Glutamat neuron), fra Striatum til den interne del af Globus Pallidus (Gpi) og pars reticulata af Substantia Nigra (SNr) (via GABA og Substans P), videre til Thalamus (via GABA) og til slut til Cortex (via Glutamat).
Det indirekte kredsløb går fra Cortex til Striatum (via Glutamat), fra Striatum til den eksterne del af Globus Pallidus (Gpe) (via GABA og Enkefalin). Herfra kan det enten gå videre til den interne del af Globus Pallidus (Gpi) og pars reticulata Substantia Nigra (SNr) (via GABA) – eller ned til Nc. subthalmicus (via GABA) og så til Gpi/SNr (via Glutamat). Herfra går det så videre til Thalamus (via GABA) og cortex (via Glutamat).
Hvis vi anvender ovenstående regneregler, så vil det direkte kredsløb altså gå igennem Glutamat (+) -> GABA/Substans P (-) -> GABA (-) -> Glutamat (+) – dvs. (+)x(-)x(-)x(+) = (+) – altså er resultatet en øget aktivitet i neuronet mod cortex.
Anvender vi det på det inddirekte kredsløb så går det altså enten via Glutamat (+) -> GABA/Enkefalin (-) -> GABA (-) -> GABA (-) -> Glutamat (+) – dvs. (+)x(-)x(-)x(-)x(+)=(-) – resultatet er altså her en nedsat aktivitet i neuronet mod cortex. Alternativt kan det gå via Glutamat (+) -> GABA/Enkefalin (-) -> GABA (-) -> Glutamat (+) -> GABA (-) -> Glutamat – dvs. (+)x(-)x(-)x(+)x(-)x(+)=(-) – resultatet er altså også her en nedsat aktivitet i neuronet mod cortex. Ligemeget hvilken vej vi vælger i det indirekte kredsløb er resultatet altså inhibitorisk.
For at opsummere, så er effekten af det direkte kredsløbet altså excitatorisk og effekten af det indirekte kredsløb er inhibitorisk.
Dopamins effekt på direkte og indirekte kredsløb
Basalgangliernes kredsløb reguleres også af Dopamin. Reguleringen er også indtegnet på ovenstående figur. Dopaminen frigives af pars compacta Substantia Nigra. Her kan det påvirke GABA-neuronerne i det direkte kredsløb via en D1 receptor (samlet betegnelse for D1 og D5 receptorer) eller det indirekte kredsløb via en D2 receptor (samlet betegnelse for D2, D3 og D4 receptorer)
Dopamin fremmer det direkte kredsløb via D1 og hæmmer det indirekte kredsløb via D2. Det direkte kredsløb var jo samlet set excitatorisk, mens det indirekte kredsløb var inhibitorisk. Derfor vil Dopamins rolle – uanset om det påvirker det direkt eller indirekte kredsløb være excitatorisk.
Dopaminerge neuroner aktiveres særligt af stimuli, der er uventede eller opleves som særligt relevante for dyrets adfærd i øjeblikket.
Dopamin frigivelsen i striatum er reguleret, så den kan frigives meget lokalt, mens der ikke frigives DA et andet sted i striatum. På denne måde spiller lokalt frigivet dopamine en rolle i at vælge et bestemt motor program via aktivering af en bestemt population af direkt pathway neuroner med D1 receptorer og hæmning af omkringliggende indirekte pathway neuroner med D2 receptorer.
Acetylcholins effekt på det direkte og indirekte kredsløb
Acetylcholin frigives også i Striatum. Dens effekt er hæmning af Dopamin – og dermed en hæmning af det direkte kredsløb eller en forstærkning af det indirekte kredsløb. Effekten er altså den modsatte af Dopamin og resultatet er nedsat stimuli i cortex.
Sygdomme
Der findes forskellige sygdomme forbundet med en forstyrrelse af basalgangliernes kredsløb. Hvis der er skader på det direkte kredsløb har man symptomer som hypokinesi – dvs. få bevægelser. Er der skader på det indirekte kredsløb har man symptomer som hyperkinesi og dyskinesi, dvs. ufrivillige, overdrevne bevægelser.
Parkinsons sygdom
Den mest kendte sygdom er nok Morbis Parkinsons som også hedder Parkinsons sygdom. I denne sker der en degenerering af neuronerne i substantia nigra pars compacta (SNc), som jo var dem der leverede dopamin til Striatum. Symptomer er hypokinesi (få bevægelser), akinesi (besvær med at initiere bevægelser), bradykinesi (langsomme bevægelser), rigiditet, hviletremor og påvirkede posturale reflekser.
Behandlingen af Parkinsons sygdom er Dopamin. Desværre kan Dopamin ikke træde over blodhjernebarrieren og derfor giver man L-DOPA i stedet. Dette er et predrug, som kan træde over blodhjernebarrieren og her omdannes til Dopamin. Omdannelsen sker ved en carboxylase der findes i neuronerne.
Desværre findes carboxylase også i resten af kroppens celler, så man får forøget Dopamin i resten af kroppen og dertil hørende symptomer. Derfor giver man det sammen med en carboxylasehæmmer (carbidopa eller benserazide), der ikke kan træde over blodhjernebarrieren. Således forhindrer man L-DOPA omdannelsen til Dopamin i kroppen – men ikke i hjernen.
Den farmakologiske behandling virker kun i de første 3-5 år efter sygdommen er brudt ud. Herefter er man nemlig resistent mod behandlingen, fordi degenereringen fortsætter. Herudover er der også bivirkninger idet en for høj dosis af L-DOPA også kan frembringe symptomer der minder om Parkinsons Sygdom.
Nyere eksperimentel behandling er også igang med at blive udviklet. Her opererer man elektroder ind i Nc. subthalmicus som får cellernes aktivitet til at falde og svarer til en hæmning af det indirekte kredsløb. Dette kaldes for Deep brain stimulation.
Huntingtons chorea
Ved Huntingtons chorea sker der en degenerering af celler i Striatum, hvor man ser et tab af GABA+Enkefalin neuroner – dvs. de neuroner der deltager i det indirekte kredsløb.
Derved får man en mindre hæmning af thalamus og dermed en øget stimulation i Cortex. Symptomer er dyskinsi (ufrivillige bevægelser). Navnet Chorea betyder også “dans” og henviser netop til disse ufrivillige bevægelser.
Det er en dominant arvelig sygdom der debuterer i 40-50 års alderen. Den skyldes CAG-repeats, som leder til dannelsen af et abnormt, toksisk protein, som ødelægger neuronerne. CAG-repeats ophobes for hver generation, og sygdommen debuterer tidligere og tidligere og har værre og værre forløb for hver generation.
Hemiballisme
Ved sygdommen Hemiballisme ses en degeneration af celler i Nc. subthalmicus. Man ser det ved apopleksi og traume og symptomerne er voldsomme, halvsidige (kontralaterale) bevægelser.
Tourettes syndrom
Ved sygdommen tourettes syndrom ses forstyrrelser i basalgangliernes samspil med præfrontalcortex, hvilket betyder at motorprogrammerne ikke kan holdes adskilt.
Man mener det muligvis hænger sammen med en dopaminerg overaktivitet. Sygdommen opstår i 5-10 års alderen. Symptomer er multiple tics (muskelkontraktioner) og vokale tics.
Sidst opdateret 31. maj 2023